
 ACCESS
Research Article
ACCESS
Research Article
Volume 2, Article ID: 2026.0008

Javier Burgos-Salcedo
jdariob@sanmateo.edu.co

1 Facultad de Ingeniería, Fundación Universitaria San Mateo; 111411 Bogotá, Colombia
2 Research Department, Corporación para la investigación y la innovación-CIINAS, 251201 Bogotá, Colombia
Received: 27 Jul 2025 Accepted: 28 Dec 2025 Available Online: 28 Dec 2025
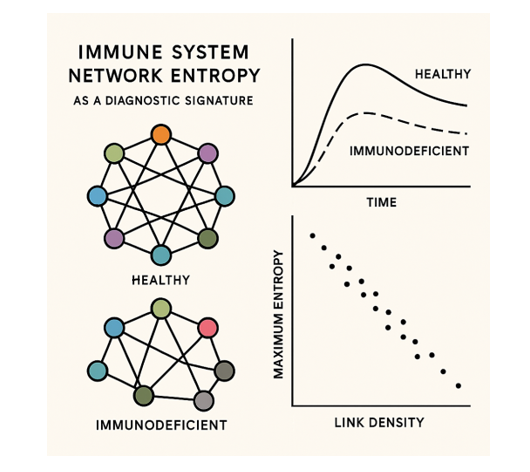
Determining the fitness status of the immune system is one of the central challenges of modern biomedical sciences, due to the relationship it has with senescence processes and aging. Currently, there is no universally valid metric that allows establishment when an individual's immune system is competent and to what degree it is competent in each pathology. In the present study, a model of complex adaptive networks is developed, whose nodes are made up of the eleven central types of immune cells, focused on the phase transitions of entropy to distinguish the optimal functioning of the immune network in contrast to what occurs in cases of immunodeficiency states. both primary and secondary. The results of the dynamic simulation indicate the presence of differentiated profiles between the immunocompetence and immunodeficiency states of the immune system and provide evidence on the state of the complex immune network and its internal connectivity, suggesting the presence of aging patterns of the immune response effector networks. This entropy-based approach offers quantitative, holistic assessment of immune status with potential applications in AI-driven diagnostic systems, disease monitoring, and personalized immunomodulatory therapies.
Disclaimer : This is not the final version of the article. Changes may occur when the manuscript is published in its final format.